Реклама
Книги по философии
Игорь Кондрашин
Диалектика Материи
(страница 25)
Вместе с тем, с определенного момента этот процесс стал протекать одновременно с началом развития фн. систем более высокого организационного уровня, фн. ячейки которых клетки стали заполнять в качестве фщ. единиц. В итоге клетка превратилась в сложное системное образование, для поддержания фн. свойств которого внутри и вне ее постоянно протекают сложные химические процессы. Постоянство процессов связано с тем, что время функционирования фщ. единиц по мере увеличения их молекулярного веса все более не совпадает со временем существования фн. ячеек структур, которые они заполняют, так как при ограниченном пространстве перемещения фщ. единиц время их существования находится в прямой зависимости от их фщ. массы. Кроме того, постоянство процессов вызвано тем, что большинство протекающих в клетке химических реакций имеет необратимый характер. Для всех этих реакций свойственны величайшая организованность и упорядоченность: каждая реакция протекает в строго определенном месте в строго определенное время в строго определенной последовательности. Молекулы ферментов расположены на мембранах митохондрий и эндоплазматической сети в том порядке, в котором идут реакции.
В клетке имеются около тысячи ферментов, с помощью которых протекают два типа реакций: синтеза и расщепления. Основным (созидающим) типом реакций следует считать реакции синтеза, в процессе которых образуются сложные молекулярные соединения, заполняющие в качестве фщ. единиц фн. ячейки подсистемных структур клетки. Так, на замену каждой отфункционировавшей молекулы белка, покинувшей ту или иную фн. ячейку, на освободившееся место поступает новая молекула белка, по структуре и химическому составу, а следовательно и своим фн. свойствам, полностью адекватная предыдущей фщ. единице. Это означает, что вновь синтезированная фщ. единица способна (или должна быть способна) принять полноценное участие в любом из характерных для данной фн. ячейки алгоритмов.
Синтезирование фщ. единиц осуществляется с помощью функционирования специальных подсистем клетки на основе кодированной генозаписи ДНК. Имеющие при этом место флуктуационные отклонения в случае их положительного эффекта обратной связью фиксируются в генозаписи и служат целям дальнейшего совершенствования данной системной структуры. При отрицательном эффекте от вновь синтезированной фщ. единицы нарушается выполнение части фн. алгоритмов и, в случае, если система не в состоянии это элиминировать, ненадлежащее функционирование соответствующей подсистемы в конечном итоге может привести к разрушению структуры данной клетки в целом. Таким путем организационное устройство клетки позволяет ей поддерживать постоянное наличие соответствующих фщ. единиц в фн. ячейках своих подсистем, что сохраняет ее структуру и чем обеспечивается способность клетки выполнять алгоритмы фн. ячеек систем более высокого порядка, куда она входит в качестве фщ. макроединицы. Все реакции биосинтеза (реакции ассимиляции) идут согласно общей теории систем с поглощением энергии движения в пространстве, которая, как бы увязая в структуре системы клетки, преобразуется в энергию связи между ее фн. ячейками.
Другой тип реакций - реакции расщепления - происходит с одновременным уменьшением энергии связи, преобразующейся в энергию движения. При реакциях диссимиляции фщ. единицы подсистем клетки, являющиеся системными образованиями более низкого порядка, отфункционировав, распадаются на фщ. единицы своего подуровня, готовые при необходимости вступить в новые синтезирующие реакции с целью формирования новых структур - фщ. единиц более высокого организационного уровня. Оба типа реакций тесно взаимосвязаны и представляют собой единый процесс, направленный на заполнение фн. ячеек структуры клетки дееспособными соответствующими фщ. единицами, что в итоге обеспечивает поддержание на должном уровне фн. свойств клетки, как единого целого.
Одним из основных и сложнейших типов синтезирующих реакций является биосинтез белков, протекающий в клетке постоянно в течение всего периода времени ее существования. В процессе функционирования клетки часть ее белков, принявших участие в каталитических реакциях, постепенно денатурируется, их структура, а следовательно и функции, нарушаются и они удаляются из своих фн. ячеек, а затем и из самой клетки. Их место в фн. ячейках занимают вновь синтезированные белковые молекулы, по своим фн. свойствам полностью идентичные освободившим им место фщ. единицам. Учитывая, что существует великое множество типов белковых молекул, механизм их синтезирования, совершенствовавшийся в течение долгого времени, в конечном итоге превратился в специализированную подсистему клетки с четким перечнем алгоритмов функционирования.
Программа синтеза белков, то есть информация об их структуре, записанная
и хранящаяся в ДНК, направляется в рибосомы с помощью информационных РНК (и-РНК),
синтезирующихся на ДНК и точно копирующих ее структуру. Каждой аминокислоте
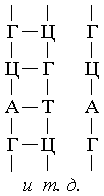
соответствует участок цепи ДНК из трех рядом стоящих нуклеотидов: А-Ц-А (цистеин),
Т-Т-Т (лизин), А-А-Ц (лейцин) и т.д. Число возможных комбинаций из 4-х нуклеотидов
по 3 равно 64, хотя используемых аминокислот всего 20. Последовательность нуклеотидов
и-РНК в точности повторяет последовательность нуклеотидов одной из цепей генозаписи,
при этом с каждого гена можно снять любое число копий РНК.
 Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности
нуклеотидов одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам,
куда поступают также и аминокислоты, доставляемые извне клетки в уже готовом
виде. Аминокислоты попадают на рибосому в сопровождении транспортных РНК (т-РНК),
состоящих в среднем из 70 - 80 нуклеотидных звеньев, в 4 - 7 местах комплементарных
друг другу. К одному из концов т-РНК присоединяется аминокислота, а в верхушке
изгиба находится триплет нуклеотидов, который соответствует по коду данной аминокислоте.
На каждую аминокислоту имеется своя т-РНК, то есть их также 20 разновидностей.
Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности
нуклеотидов одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам,
куда поступают также и аминокислоты, доставляемые извне клетки в уже готовом
виде. Аминокислоты попадают на рибосому в сопровождении транспортных РНК (т-РНК),
состоящих в среднем из 70 - 80 нуклеотидных звеньев, в 4 - 7 местах комплементарных
друг другу. К одному из концов т-РНК присоединяется аминокислота, а в верхушке
изгиба находится триплет нуклеотидов, который соответствует по коду данной аминокислоте.
На каждую аминокислоту имеется своя т-РНК, то есть их также 20 разновидностей.
Синтез белков и нуклеиновых кислот происходит на основе реакций матричного синтеза. Этим обеспечивается придание вновь образованным соединениям фн. свойств заменяемых фщ. единиц. Новые молекулы синтезируются в точном соответствии с планом, заложенным в структуре уже существующих молекул. Поэтому в этих реакциях обеспечивается точная, строго специфичная последовательность мономерных звеньев в синтезируемых полимерах. Здесь происходит направленное стягивание мономеров в определенное место клетки - в фн. ячейки вновь образуемого полимера, при этом расположение самих фн. ячеек предопределяется структурной организацией копируемой матрицы. Роль матрицы в матричных реакциях играют макромолекулы нуклеиновых кислот ДНК и РНК. Мономерные молекулы (нуклеотиды или аминокислоты) в соответствии с принципом комплементарности располагаются и фиксируются на матрице в строго определенном, заданном порядке. Затем происходит "сшивание" мономерных звеньев в полимерную цепь, и готовый полимер сбрасывается с матрицы. После этого матрица готова к сборке новой полимерной молекулы. При помощи матричного типа реакций осуществляется воспроизведение однотипных соединений - фщ. единиц данной системы. Потребность воспроизведения однотипных фщ. единиц прослеживается на всех уровнях организации Материи и является одной из главных закономерностей общей теории систем.
Информация о структуре белка, записанная в и-РНК в виде последовательности нуклеотидов, переносится далее в виде последовательности аминокислот в синтезируемую полипептидную цепь, то есть происходит процесс "трансляции". По мере сборки белковой молекулы рибосома ползет по и-РНК, за ней вторая, третья и т.д. Каждая из них синтезирует один и тот же белок, запрограммированный на данной и-РНК. Когда рибосома пройдет и-РНК от конца до конца - синтез белка окончен. После этого рибосома идет на другую и-РНК, а белок по эндоплазматической сети направляется в соответствующую его фн. свойствам свободную фн. ячейку, которую он заполняет в качестве фщ. единицы.
Синтез белков в клетке происходит непрерывно. Все рибосомы, помещающиеся одновременно на одной и-РНК, объединяются в полирибосому. Рибосома работает вдоль и-РНК "шажками": триплет за триплетом РНК находится в контакте с нею. Для сшивания полипептидной цепи в рибосоме имеется белок - синтетаза. Молекулы т-РНК, проходя через рибосому, задевают своим кодовым концом место контакта рибосомы с и-РНК. Если кодовый триплет т-РНК окажется комплементарным к триплету и-РНК, аминокислота, доставленная т-РНК, переходит из ее фн. ячейки в фн. ячейку синтезируемой молекулы белка, таким образом становясь фщ. единицей ее структуры. Этим обеспечивается наиважнейшее правило построения фн. систем - помещение данной фщ. единицы в строго соответствующую ей фн. ячейку или, наоборот, заполнение фн. ячейки строго соответстующей ей фщ. единицей. Поэтому имеющийся в любой клетке механизм синтезирования белков обеспечивает стопроцентную гарантию того, что переносимая т-РНК данная аминокислота попадет только в соответствующую ей фн. ячейку структуры белка или, наоборот, того, что в подошедшую в рибосоме незаполненную очередную фн. ячейку синтезируемого белка попадет только соответствующая ей по фн. свойствам фщ. единица - требуемая аминокислота.